

There is a mechanism of macrolide resistance in Staphylococcus spp. which also affects the lincosamides and type B streptogramins characterizing the so-called MLSB resistance, whose expression can be constitutive (cMLSB) or inducible (iMLSB) and is encoded mainly by ermA and ermC genes. The cMLSB resistance is easily detected by susceptibility testing used in the laboratory routine, but iMLSB resistance is not. Therapy with clindamycin in cases of infection with isolated iMLSB resistance may fail.
ObjectiveTo characterize the phenotypic (occurrence of cMLSB and iMLSB phenotypes) and molecular (occurrence of ermA and ermC genes) profiles of MLSB resistance of clinical isolates of susceptible and methicillin-resistant Staphylococcus aureus and CNS (coagulase-negative Staphylococcus) from patients of a university hospital, in Pernambuco.
MethodsThe antimicrobial susceptibility of 103 isolates was determined by the disk diffusion technique in Mueller–Hinton agar followed by oxacillin screening. The iMLSB phenotype was detected by D test. Isolates with cMLSB and iMLSB phenotypes were subjected to polymerase chain reaction (PCR) for the detection of ermA and ermC genes.
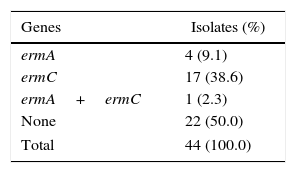
ResultsThe cMLSB and iMLSB phenotypes were respectively identified in 39 (37.9%) and five (4.9%) isolates. The iMLSB phenotype was found only in four (10.8%) methicillin-susceptible S. aureus and one (4.5%) methicillin-resistant S. aureus. In the 44 isolates subjected to PCR, four (9.1%) only ermA gene was detected, a lower frequency when compared to only ermC 17 (38.6%) gene and to one (2.3%) isolate presenting both genes.
ConclusionIn the Staphylococcus spp. analyzed, the ermC gene was found more often than the ermA, although the iMLSB phenotype had been less frequent than the cMLSB. It was important to perform the D test for its detection to guide therapeutic approaches.
The increasing prevalence of methicillin resistance in Staphylococcus spp. is a growing problem. This has renewed interest regarding the use of macrolide, lincosamides, and type B streptogramin antimicrobials for the treatment of staphylococcal infections. Clindamycin, a lincosamide, represents a common choice for some of these infections, particularly for infections of the skin and soft tissues, and an alternative in case of intolerance to penicillin or methicillin resistance.1–3 In Staphylococcus spp., one of the resistance mechanisms consists of ribosomal target modification, affecting macrolides, lincosamides, and type B streptogramins characterizes the so-called MLSB resistance. Its expression can be constitutive (cMLSB) or inducible (iMLSB) and is encoded by ermA (erythromycin ribosome methylase) and ermC genes, which are the main determinants of resistance in staphylococci.2,4,5
It is important to know the type of MLSB resistance for establishing adequate therapy, since Staphylococcus spp. with constitutive resistance present in vitro resistance to all macrolides, lincosamides, and type B streptogramins. In addition, in vivo therapy with clindamycin for Staphylococcus spp. infection with inducible resistance can select constitutive erm mutants, resulting in treatment failure.3,4,6,7 It is noteworthy that cMLSB resistance is easily detected by susceptibility testing used in the laboratory routine, while iMLSB resistance is not. Using these detection methods, Staphylococcus spp. with inducible resistance has in vitro resistance to erythromycin and susceptibility to clindamycin.5,6,8,9
To detect the inducible clindamycin resistance in Staphylococcus spp., one of the tests recommended by the CLSI (Clinical and Laboratory Standards Institute) is the double-disk diffusion test (D Test) and when the isolated present such resistance, the CLSI recommends reporting them as resistant to clindamycin.10 Then, data on the antimicrobial susceptibility are important in the choice of therapy against infections, but false susceptibility results can be obtained if the isolates are not subjected to tests that detect inducible resistance to clindamycin.11
Studies carried out in two Brazilian states with clinical isolates of Staphylococcus spp. reported the cMLSB phenotype as the most frequent.12,13 Coutinho et al.13 have also evaluated the occurrence of the erm genes among the isolates analyzed. However, the frequency of cMLSB and iMLSB resistance varies among different hospitals and there are other resistance mechanisms that confer resistance to only one or two classes of the MLSB complex.14,15
The objective of this study was to characterize the phenotypic (occurrence of cMLSB and iMLSB phenotypes) and molecular (occurrence of ermA and ermC genes) profiles of MLSB resistance of clinical isolates of susceptible and methicillin-resistant Staphylococcus aureus and CNS (coagulase-negative Staphylococcus) from patients of a university hospital in Pernambuco, Brazil. Obtaining local data relating to resistance, may be helpful in guiding therapeutic approaches.
Materials and methodsClinical isolatesA total of 103 clinical isolates were gathered from various samples from patients infected with S. aureus or SCN of a university hospital of Pernambuco, Brazil, during the year 2012 and were stored in glycerol (25%) at −20°C. To verify the purity, the colonies were inoculated into Brain Heart Infusion broth (BHI) and after incubation at 37°C for 48h were plated on blood agar.
Antimicrobial susceptibility profileThe antibiogram was performed by disk diffusion technique in Mueller–Hinton agar, using antibiotic clindamycin 2μg, erythromycin 15μg, cefoxitin 30μg, and oxacillin 1μg. The results were interpreted according to the standards determined by CLSI.10
Screening for oxacillin resistanceIsolates with resistance or intermediate resistance to oxacillin and/or cefoxitin were submitted to oxacillin screening, as proposed by Rabelo et al.16
D testS. aureus and SCN isolates with resistance to erythromycin and susceptibility or intermediate resistance to clindamycin in the antibiogram were selected. For the execution of this test a disk of 2μg of clindamycin was placed at a distance of 15mm–26mm from the edge of a disk of 15μg of erythromycin in a plate containing Mueller–Hinton agar sown in the same way as it was for the antibiogram. After incubation at 35°C for 16–18h, isolates that showed no flattening of the inhibition zone around the clindamycin disk were reported as susceptible to clindamycin (negative D test) and isolates that showed flattening of the inhibition zone around the clindamycin disk adjacent to erythromycin disk (“D” zone) indicated inducible clindamycin resistance (positive D test).10
Extraction of total DNATo examine the presence of ermA and emrC genes, the rapid extraction of the total DNA of isolates that showed phenotypes MLSBc and MLSBi was performed by a modified technique of thermal lysis directly from the colony, according to Hu et al.,17 after inoculation of a colony of each isolate into 5mL of BHI and incubated at 37°C for 24h.
Polymerase chain reaction (PCR) conditionsPCR was performed using the primers described by Lina et al.18 for ermA and ermC genes. For the detection of ermA gene, each amplification reaction was prepared in a final volume of 25μL for each tube and includes: 1μL (40ng) of total DNA, 1μL (20pmol) of each primer, 0.6μL of deoxyribonucleotide triphosphate (dNTP) (8mM), 5.0μL of buffer (5×), 1.5μL of MgCl2 (25mM), 0.4μL of Taq DNA polymerase (5U). For the detection of ermC gene, each amplification reaction was prepared in the same manner as for ermA gene, except in relation to the quantity of Taq DNA polymerase (5U), which in this reaction was 0.3μL. Amplification reactions were performed in the thermocycler at the following conditions: 30 cycles of 1min at 94°C in the denaturing step, 30s at 49°C in the annealing step and 30s at 72°C in the extension step, according to the conditions described by França et al.19 with modification. In all PCR a negative control was included, corresponding to a tube containing all components of the mixture to which template DNA was not added. Identified positive isolates for the ermA and ermC genes were used as positive controls. The PCR products were subjected to electrophoresis on 1.5% agarose gel in 0.5× TBE buffer. These products were stained with blue, visualized on an ultraviolet transilluminator and photodocumented.
Sequencing of ermA and ermC genesA PCR product positive for the ermA and ermC genes were purified through the kit Wizard® SV Gel and PCR Clean-Up System (Promega), according to manufacturer's protocol and then, quantified by spectrophotometry. Thereafter they were submitted to sequencing and analysis of the results was performed through the softwares Chromas Lite 2.1.1, Basic Local Alignment Search Tool (BLAST), and Expert Protein Analysis System (ExPASy). The analyzed sequences of ermA and ermC genes were deposited in the GenBank with the following access numbers, respectively: KT599443 and KU232395.
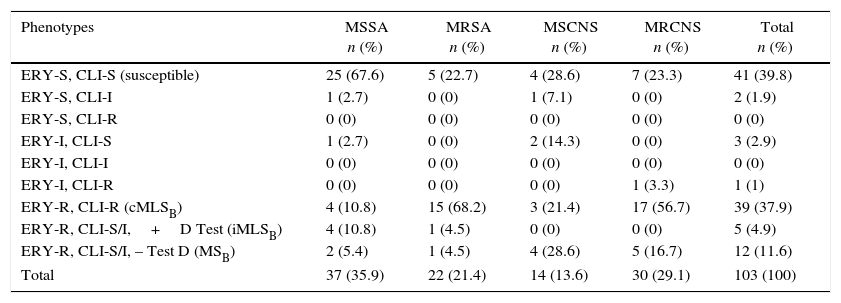
ResultsFifty-nine (57.3%) clinical isolates of S. aureus and 44 (42.7%) of SCN were analyzed, totaling 103 isolates. Of these, 37 (35.9%), 22 (21.4%), 14 (13.6%), and 30 (29.1%) were classified as methicillin-susceptible S. aureus (MSSA), methicillin-resistant S. aureus (MRSA), methicillin-susceptible coagulase-negative Staphylococcus (MSCNS) and methicillin-resistant coagulase-negative Staphylococcus (MRCNS), respectively (Table 1).
Susceptibility profile to erythromycin and clindamycin of susceptible and methicillin-resistant S. aureus and CNS.
| Phenotypes | MSSA n (%) | MRSA n (%) | MSCNS n (%) | MRCNS n (%) | Total n (%) |
|---|---|---|---|---|---|
| ERY-S, CLI-S (susceptible) | 25 (67.6) | 5 (22.7) | 4 (28.6) | 7 (23.3) | 41 (39.8) |
| ERY-S, CLI-I | 1 (2.7) | 0 (0) | 1 (7.1) | 0 (0) | 2 (1.9) |
| ERY-S, CLI-R | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| ERY-I, CLI-S | 1 (2.7) | 0 (0) | 2 (14.3) | 0 (0) | 3 (2.9) |
| ERY-I, CLI-I | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| ERY-I, CLI-R | 0 (0) | 0 (0) | 0 (0) | 1 (3.3) | 1 (1) |
| ERY-R, CLI-R (cMLSB) | 4 (10.8) | 15 (68.2) | 3 (21.4) | 17 (56.7) | 39 (37.9) |
| ERY-R, CLI-S/I,+D Test (iMLSB) | 4 (10.8) | 1 (4.5) | 0 (0) | 0 (0) | 5 (4.9) |
| ERY-R, CLI-S/I, – Test D (MSB) | 2 (5.4) | 1 (4.5) | 4 (28.6) | 5 (16.7) | 12 (11.6) |
| Total | 37 (35.9) | 22 (21.4) | 14 (13.6) | 30 (29.1) | 103 (100) |
ERY, erythromycin; CLI, clindamycin; S, susceptible; I, intermediary; R, resistant; MSSA, methicillin-susceptible Staphylococcus aureus; MRSA, methicillin-resistant S. aureus; MSCNS, methicillin-susceptible coagulase-negative Staphylococcus; MRCNS, methicillin-resistant coagulase-negative Staphylococcus; MLSB, macrolides, lincosamides and type B streptogramins; cMLSB, phenotype of constitutive MLSB resistance; iMLSB, phenotype of inducible MLSB resistance; MSB, phenotype of resistance to macrolides and type B streptogramins.
The susceptible phenotype (ERY-S, CLI-S) was detected in 25 (67.6%), five (22.7%), four (28.6%), and seven (23.3%) of MSSA, MRSA, MSCNS and MRCNS, respectively (Table 1). The phenotype cMLSB (ERY-R, CLI-R) was detected respectively, in four (10.8%) 15 (68.2%), three (21.4%), and 17 (56.7%) of MSSA, MRSA, MSCNS and MRCNS. The iMLSB phenotype (ERY-R, CLI-S, positive D test) (Fig. 1) was found in only four (10.8%) of the MSSA and in one (4.5%) of the MRSA. The MSB phenotype (ERY-R, CLI-S, negative D test) was detected in two (5.4%), one (4.5%), four (28.6%), and five (16.7%) of MSSA, MRSA, MSCNS, and MRCNS, respectively (Table 1).
.")
For the identification of the ermA and ermC genes, 44 isolates with cMLSB and iMLSB phenotypes underwent PCR. Four (9.1%) isolates showed only ermA gene, 17 (38.6%) had only ermC gene, one (2.3%) had both genes, and 22 (50%) did not have the referred genes (Table 2).
DiscussionIn this study, the frequency of MSSA (25; 67.6%) was greater than MRSA (five; 22.7%) and of MSCNS (four; 28.6%) was lower than MRCNS (seven; 23.3%). In a study conducted in India, 313 isolates of Staphylococcus spp. there were 83 (64.84%) MSSA and 45 (35.15%) MRSA, in line to the present study; in contrast, there were 124 (67.02%) MSCNS and 61 (32.97%) MRCNS.5 A total of 1687 isolates of Staphylococcus spp. investigated in Turkey consisted of 419 (24.8%) MSSA and 464 (27.5%) MRSA, unlike this study, and the rest of the isolates, similar to this study, consisted of 196 (11.6%) MSCNS and 608 (36.1%) MRCNS.11
The susceptible phenotype (ERY-S, CLI-S) found in 41 (39.8%) isolates in this study prevailed among the isolates tested. This phenotype was also prevalent in studies carried out in India, Libya, and Turkey where the referred phenotype was detected in 192 (51.5%), 87 (54.7%), and 688 (40.8%) isolates, respectively.11,20,21
In this study, the 39 (37.9%) cMLSB phenotype isolates prevailed over the five (4.9%) iMLSB phenotype and over the 12 (11.6%) MSB phenotype resistant to macrolides and type B streptogramins, which is consistent with other studies conducted in Brazil.12,13,22 One of the studies was performed in São Paulo revealed 37 (25.17%) and nine (6.12%) isolates with cMLSB and iMLSB phenotypes, respectively, while the MSB was not detected.12 Other studies were conducted in the state of Rio Grande do Sul; one of them identified 71 (46.7%), five (3.3%), and five (3.3%) isolates with cMLSB, iMLSB, and MSB phenotypes, respectively.13 The other study detected 25 (17.9%), 11 (7.9%), and two (1.4%) isolates with cMLSB, iMLSB, and MSB phenotypes, respectively.22
In this study the cMLSB phenotype prevailed among the isolates resistant to methicillin, which is consistent with studies conducted in other countries3,5,11,15,21 as well in Brazil.12,22
The MSB phenotype was more frequent (12; 11.6%) than the iMLSB (five; 4.9%) in this study. Diverging from this finding, the study done by Kumar et al.8 in India found the same frequency (33; 16.9%) of MSB and iMLSB phenotypes among the S. aureus isolates analyzed. Merino-Díaz et al.23 in Spain identified the msrA gene in all Staphylococcus spp. isolates with MSB phenotype. In Staphylococcus spp. this gene is responsible for an efflux mechanism which confers resistance to macrolides and type B streptogramins, but not to clindamycin.5,24
Only S. aureus showed the iMLSB phenotype and its frequency was higher among the MSSA (four; 10.8%) than among the MRSA (one; 4.5%) in this study. Astudy conducted in Chicago detected the iMLSB phenotype in 59 (20%) MSSA isolates and in 14 (7%) MRSA isolates in one hospital, and the iMLSB phenotype in 94 (19%) and in 30 (12%) MSSA and MRSA isolates, respectively, in another hospital.25 In a study conducted in Turkey, eight (5.8%) MSSA isolates and two (1.7%) MRSA isolates presented the iMLSB phenotype.26 In the study conducted in São Paulo, the iMLSB phenotype was observed in seven (6.73%) MSSA and in two (4.65%) MRSA isolates.12 These data from previously reported studies are similar to the findings of the present study. However, there are other studies carried out abroad, in which the iMLSB phenotype was prevalent among MRSA isolates.3,8,15,27,28
Relevant data for antimicrobial susceptibility are significant in establishing an appropriate therapy; therefore, the availability of the D test results are important.20,25 Reporting an isolated Staphylococcus spp. as susceptible to clindamycin without verifying if it presents inducible resistance, may lead to inadequate therapy with this drug. In contrast, a negative result for inducible resistance to clindamycin, confirms the susceptibility of this antimicrobial, providing a very good therapeutic option.3,8
In this study, the ermA and ermC genes were detected individually in four (9.1%) and 17 (38.6%) isolates with MLSB resistance phenotypes, respectively. In some studies, contrary to this, the ermA gene was identified more frequently than the ermC gene, as in a study conducted in Iran, in which the ermA and ermC genes were found in 76 (60.3%) and 69 (54.8%) S. aureus isolates, respectively.29 Schmitz et al.,30 when analyzing S. aureus isolates from 24 European university hospitals detected the ermA gene in 571 (67%) isolates and the ermC in 192 (23%). In Korea, Jung et al.31 identified among 280 S. aureus isolates, the ermA gene in 250 and the ermC in 14 isolates. In Iran, Moosavian et al.32 detected the ermA and ermC genes, respectively in 41.1% and 17.7% of the S. aureus isolates studied.
In other studies, consistent with this, the ermC gene was predominant relatively to the ermA gene, as in a study from Spain, in which the ermC gene was more frequently detected, in both S. aureus and SCN isolates with constitutive and inducible phenotypes.23 In Greece, Spiliopoulou et al.33 identified the ermA and ermC genes in 22% and 70% of S. aureus isolates, respectively. Vallianou et al.34 have reported that the ermC gene was found more frequently among S. aureus and SCN isolates of a university hospital also in Greece.
In this study, one (2.3%) isolate presented both ermA and ermC genes. The association of these genes in Staphylococcus spp. isolates,29,35,36 was also observed in other studies.
The ermB gene was not investigated, because it is usually detected in Staphylococcus spp. isolates of animal origin13,23,30 and it is spread mainly between streptococci and enterococci.4 In Rio Grande do Sul, Coutinho et al.13 reported low frequency of the ermB gene. They found that among 152 Staphylococcus spp. isolates analyzed, 77 had one or more erm gene, the ermA, ermC, and ermB genes were found respectively at 49, 29, and three isolates and, the combination of these genes was found in four isolates,13 data which differed from this study. However, in a work developed in Texas, with S. aureus isolates of pediatric origin, among the 67 isolates evaluated, the ermB gene was identified in 31 of these, and the ermA and the ermC genes were detected in 12 and 24 isolates, respectively.37
Then it was possible to observe the frequency variation of the phenotypes of the MLSB resistance and of the erm genes amongst hospitals and geographic regions, already reported by other authors.13,15,21,29 Therefore, the importance of determining these frequencies in specific location is highlighted.13,21
The good correlation between the phenotypic and genotypic methods allows us to infer the mechanism of resistance to erythromycin and clindamycin, to establish the most appropriate antimicrobial treatment and to appreciate the epidemiological differences in their distribution.23
ConclusionThe ermA gene was less frequent than the ermC gene among Staphylococcus spp. isolates with cMLSB and iMLSB phenotypes. Despite the lower frequency of iMLSB than cMLSB and MSB phenotypes, it is important to perform the D test in order to identify it and guide therapeutic procedures. As the phenotypic and molecular data about a particular mechanism of antimicrobial resistance vary between hospitals and geographic regions, obtaining local data is useful, since it can be helpful to emphasize the importance of the implementation of procedures that aim at controlling the spread of this mechanism, in the hospital where the study was conducted.
Conflicts of interestThe authors declare no conflicts of interest.
This study was supported by the National Counsel of Technological and Scientific Development (CNPq).
