
Hypothyroidism due to Hashimoto's thyroiditis (HT) is the commonest autoimmune endocrine illness in which antibodies against thyroid organ result in inflammation. The disease has a complex etiology that involves genetic and environmental influences. Viral infections may be involved in triggering of the disease as their molecular mimicry enhance autoimmune responses. Human herpesvirus-6 (HHV-6) is recognized for its contribution to some autoimmune diseases.
ObjectiveIn the current study, the prevalence of HHV-6 active infection in patients with HT and with non-autoimmune thyroid disorders were compared with patients with euthyroidism. In addition, a correlation between presence of HHV-6 infections and HT was investigated.
MethodsA total of 151 patients with clinically and laboratory confirmed HT, 59 patients with non-autoimmune thyroid disorders, and 32 patients with normal thyroid function were included in the study. For further confirmation of HT disease, all the precipitants were tested for anti-thyroid peroxidase (TPO), and anti-thyroglobulin (TG) antibodies. For detection of both HHV-6 types A and B, nested PCR and restriction enzyme digestion were used. HHV-6 DNA positive samples were further investigated by DNA sequencing analysis.
ResultsHHV-6A DNA was found in serum sample of 57 out of 151 patients (38%) with HT, which was significantly more often than in patients with non-autoimmune thyroid disorders (p=0.001). However, HHV-6 DNA was not detected in serum samples of euthyroid subjects.
ConclusionsThe results support a possible role for active HHV-6A infection, demonstrated by the presence of HHV-6 DNA in sera, in the development of HT.
Hashimoto's thyroiditis (HT) is a common autoimmune disease in which the immune system attacks the thyroid causing inflammation and progressive destruction of the gland. It is a common cause of hypothyroidism with a potential complication of thyroid lymphoma.1,2
Several factors are thought to be involved in the etiology of the disease including genetic and environmental factors.3 Viral infection may play a role in triggering autoimmunity through molecular mimicry.4
Recently, human herpesviruses 6 type A and type 6B were recognized as two distinct viruses with different biological features and disease associations.5 HHV-6B is associated with roseola infantum, a benign childhood febrile disease, which can be complicated by febrile seizures. In addition, it is the major cause of encephalitis in transplant patients.6
While HHV-6B is common and acquired by over 70% of children by age three,7 HHV-6A is acquired later in life and the prevalence is unknown. In preliminary reports, HHV-6A has been associated with Alzheimer's disease8 and unexplained infertility.9 HHV-6A/B have also been proposed as triggers for a number of autoimmune disorders, including multiple sclerosis (MS),10,11 autoimmune connective tissue diseases,12 and HT.13 Seropositivity for HHV-6A/B is over 90% in many studies.14
HHV-6A and HHV-6B infect T cells, monocytes–macrophages, epithelial cells, and central nervous system cells.15 It has been shown that CD46, a complement regulatory protein, is the only cellular receptor for HHV-6A which is a surface protein presented in all cell types,5,16,17 whereas HHV-6B uses CD134, a member of the TNF-receptor, that is expressed mainly on activated T lymphocytes.18 The HHV-6A uses envelop gH/gL/gQ1/gQ2 glycoprotein complex to attach cellular CD46 receptor, while the HHV-6B rarely binds to CD46 with the same glycoproteins as HHV-6A.16
A recent study suggests a potential link between HHV-6 infection of thyroid cells and the progress or activating of HT disease.13,19 In addition, available data on in vitro infection of thyrocytes with HHV-6A indicate modulation of miRNAs which are involved in the disease development. These alterations were not seen in thyrocytes infected with HHV-6B.19
The presence of HHV-6B and HHV-6A DNA in the plasma sample both indicates active infection, though HHV-6B is more likely to go into latency in peripheral blood mononuclear cells (PBMCs). Although HHV-6B DNA is detected in PBMCs of both patients and healthy individuals, analysis of serum samples indicated that none of the healthy blood donors were positive for HHV-6 DNA.20 Considering that a possible relationship exists between infection with human herpesvirus 6 (HHV-6) and autoimmune diseases, the purpose of this study was to determine the prevalence of HHV-6A/B DNA in serum of patients with HT, non-autoimmune thyroid disorders, and patients with normal thyroid gland function (euthyroid) to identify if HHV-6A/B active infection may be a possible risk factor for triggering HT disease.
Material and methodsPatients and clinical samplesThyroiditis were classified based on the American Thyroid Association definition. Based on clinical, laboratory, and ultrasonography findings, 151 patients with a definite diagnosis of HT, 59 patients with non-autoimmune thyroid disorders, and 32 subjects with normal thyroid gland function were included in the study. To confirm the results, thyroid function tests were repeated prior to taking any drug that could possibly affect thyroid function. None of them had concomitant diseases.
The patients were initially diagnosed with thyroiditis in routine clinical practice by general practitioners and referred to the endocrinology clinic of the University Hospital for further investigation. The study was approved by the University Ethics Committee. Informed consent was collected from all participants before sample collection.
Blood samples were collected in serum gel separator tubes and were then centrifuged to separate the serum and stored in a sterile tube at −20°C until use.
Thyroid function measurements and antibody assaysSerum TSH (normal range, 0.3–5.3μIU/mL), free T3 (normal range, 1.9–5.7pmol/L), free T4 (normal range, 10–22pmol/L), thyroid peroxidase antibodies (anti-TPO normal range, <35IU/mL), and thyroglobulin antibodies (Tg Ab<115IU/mL) were measured in all subjects by using an immuno-chemiluminescence technique (Cobas E411 system). Anti-HHV-6 antibody in the serum samples was determined by enzyme immunoassay according to the manufacturer's instruction (Biotin, Dublin, Ireland) as described before.11 An index value for HHV-6 antibody in serum samples for both patient and control samples was calculated as suggested by the manufacturer (Biotin).
DNA isolationTotal DNA was isolated from the serum samples with the use of DNeasy Blood & Tissue Kit (Qiagen, Germany) according to the manufacturer's protocol. The DNA was then re-suspended in a final elution buffer volume of 50μL and stored at −20°C until further analysis.
Detection of HHV-6 DNA in serum samplesThe presence of HHV-6 DNA in serum samples was detected by nested PCR with primers designed to specifically targeting U67 gene in a region, a conserved between HHV-6A and HHV-6B. For the amplification of HHV-6 DNA, the outer primers (PF1: 5′GCTAGAACGTATTTGCTG3′; PR1: 5′ACAACTGTCTGACTGGCA3′) which amplify a fragment of 252 base pairs (bp) and the inner primers (PF2: 5′CTCAAGATCAACAAGTTG3′; PR2: 5′TCACGCACATCGGTATAT3′) which amplify a fragment of 154bp were used. Primer-BLAST was performed to check the specificity of the primers (https://www.ncbi.nlm.nih.gov/tools/primer-blast/).
Nested PCRFor the amplification of target genes, PCR was run in two separate steps. To amplify a 154bp DNA fragment of the major capsid protein (MCP) of HHV-6A and HHV-6B genotype genes, the first round of PCR amplification was carried out with 2μL of extracted DNA from serum samples, 1× PCR master mix, and 0.5μM of each outer primers in a final volume of 20μL. To construct the plasmid control for HHV-6A and HHV-6B, the PCR product fragments of HHV-6A and HHV-6B isolated from clinical samples and confirmed by DNA sequencing, U67 fragments were amplified as follows: initial denaturation at 95°C for 5min, 35 cycles of 95°C for 30s, 50°C for 30s, and 72°C for 30s, followed by a final extension at 72°C for 5min.
The second round of PCR was performed as mentioned for the first round, except that one microliter of the first PCR product was added to the second PCR reaction mixture. The PCR products were analyzed on 2% agarose gel electrophoresis and stained with GelRed TM (Biotium, UK), and visualized under UV light.
To test for possible plasmid contamination, PCR reaction was performed using T7 universal primer of pTZ57R/T vector 5′TAATACGACTCACTATAGGG3′ and PF1 primer of HHV-6 5′GCTAGAACGTATTTGCTG3′ that amplify a 320bp fragment including a portion of HHV-6 and T7 promoter sequences. In addition, extensive precautions were used to prevent PCR contamination. To ensure efficient DNA extraction and PCR amplification, a 177-bp fragment of the human β-actin gene was amplified together with the samples using 5′ATCGTGCGTGACATTAAGGAG3′ and 5′GAAGGAAGGCTGGAAGAGTG3′ primers.
The sensitivity of nested PCR assaysTo evaluate the sensitivity of HHV-6 PCR detection, a set of serum sample negative for HHV-6A and HHV-6B DNA were spiked with known 10-fold serial dilutions of HHV-6 A and HHV-6B DNA in a separate reaction. After DNA extraction, two stages of consecutive amplification were used for each virus separately. The product of the first-round amplification (252bp) was used as targets for the second round of nested PCR (154bp) amplification. The last positive dilution of the first-round PCR product (252bp) and dilutions which in the first round PCR failed to yield a PCR product were used as targets for the determination of the second round of nested PCR sensitivity test (154bp). The amplified products were then analyzed by gel electrophoresis and GelRedTM (Biotium, USA) post-staining.
Sequence analysisTo investigate the specificity of PCR, the amplified products were subjected to nucleotide sequencing analysis. To verify the accuracy of sequence determination, each nested PCR product was gel purified and cloned into pTZ57R/T plasmid vector. Sequence analysis was performed (150–200ng/μL) in both forward and reverse directions using specific primers by ABI 3730XL DNA analyzer (Bioneer, Inc.).
Restriction fragment analysis for HHV-6A/B determinationA 154bp fragment of the U67 gene was amplified. This fragment includes an XhoI restriction site in the HHV-6A genome absent in type B. XhoI digestion was performed on the PCR product, producing two fragments of 105 and 49bp for HHV-6A. The procedure was performed according to the manufacturer's instruction (Thermo scientific).
Statistical analysisStatistical analysis was performed using SPSS software version 18 (SPSS Inc., Chicago, IL, USA). Pearson Chi-Square and Likelihood Ratio were used to determine any relation between the variables. Two-tailed p-values<0.05 were considered as statistically significant.
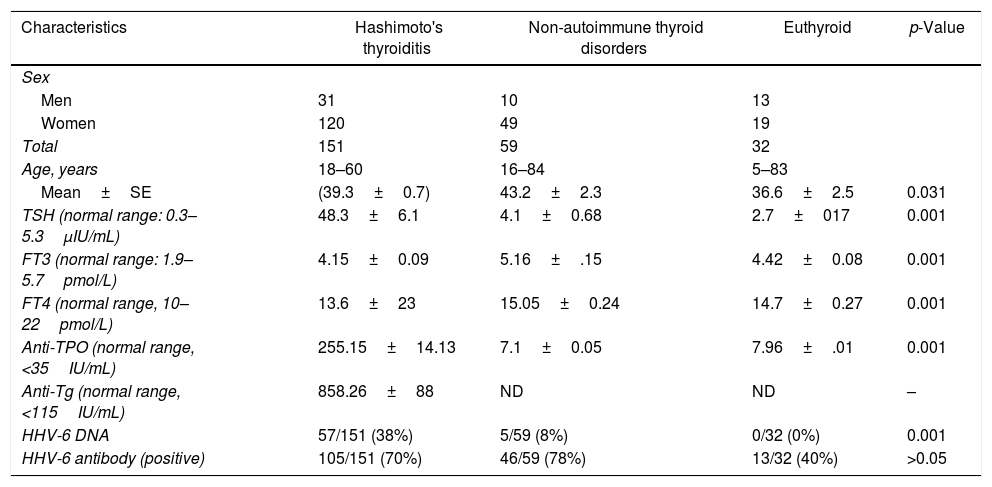
ResultsGeneral characteristicsOverall, 242 patients were enrolled in the study. The demographic characteristics of patients and the results of thyroid function tests are shown in Table 1.
Thyroid function tests, antibody assays, and HHV-6 DNA detection in study groups.
| Characteristics | Hashimoto's thyroiditis | Non-autoimmune thyroid disorders | Euthyroid | p-Value |
|---|---|---|---|---|
| Sex | ||||
| Men | 31 | 10 | 13 | |
| Women | 120 | 49 | 19 | |
| Total | 151 | 59 | 32 | |
| Age, years | 18–60 | 16–84 | 5–83 | |
| Mean±SE | (39.3±0.7) | 43.2±2.3 | 36.6±2.5 | 0.031 |
| TSH (normal range: 0.3–5.3μIU/mL) | 48.3±6.1 | 4.1±0.68 | 2.7±017 | 0.001 |
| FT3 (normal range: 1.9–5.7pmol/L) | 4.15±0.09 | 5.16±.15 | 4.42±0.08 | 0.001 |
| FT4 (normal range, 10–22pmol/L) | 13.6±23 | 15.05±0.24 | 14.7±0.27 | 0.001 |
| Anti-TPO (normal range, <35IU/mL) | 255.15±14.13 | 7.1±0.05 | 7.96±.01 | 0.001 |
| Anti-Tg (normal range, <115IU/mL) | 858.26±88 | ND | ND | – |
| HHV-6 DNA | 57/151 (38%) | 5/59 (8%) | 0/32 (0%) | 0.001 |
| HHV-6 antibody (positive) | 105/151 (70%) | 46/59 (78%) | 13/32 (40%) | >0.05 |
TSH, thyroid stimulating hormone; FT3, free tri-iodothyronine; FT4, free thyroxin; anti-TPO, thyroid peroxidase antibody; anti-Tg, anti-thyroglobulin; ND, not done.
For the whole study group, there was a significant difference between age and TSH levels of participants (p<0.05). In addition, a significant correlation between age and the levels of FT3, FT4, anti-TPO, and anti-Tg (p<0.01) was found.
Anti-IgG in patients with Hashimoto thyroiditis and in control groupsAnti-HHV-6 IgG antibody was detected in 69.5% of patients with HT, in 77.8%, patients with non-autoimmune thyroid disorders, and in 40.6% of euthyroid subjects. Based on anti-HHV-6 antibody detection, there was a significant difference between euthyroid subjects and patients suffering from thyroid dysfunctions (p<0.001).
HHV-6 DNA in serum samplesSerum samples collected from 151 HT patients and 59 patients with non-autoimmune thyroid disorders as well as 32 euthyroid patients were analyzed for the presence of HHV-6 DNA by nested PCR using primers specific for a highly conserved sequence of U67 gene corresponding to the major capsid protein gene of HHV-6A/B. When evaluated by gel electrophoresis and GelRed staining, the lower limits of detection using nested PCR assay for HHV-6A/B DNA were determined to be five copies/reaction of HHV-6A or HHV-6B DNA.
To ensure impartiality, and avoid errors arising from bias, all the samples were analyzed in double-blind tests. HHV-6 DNA was detected in the serum sample of 57 out of 151 (38%) patients with HT, in contrast to five out of 59 (8%) patients with non-autoimmune thyroid disorders and 0/32 (0%) in euthyroid subjects. The prevalence of HHV-6 DNA in HT patients compared to that of the two other control groups was significantly different (p=0.001). Distribution of HHV-6 DNA in serum samples of the patients is shown in Table 1.
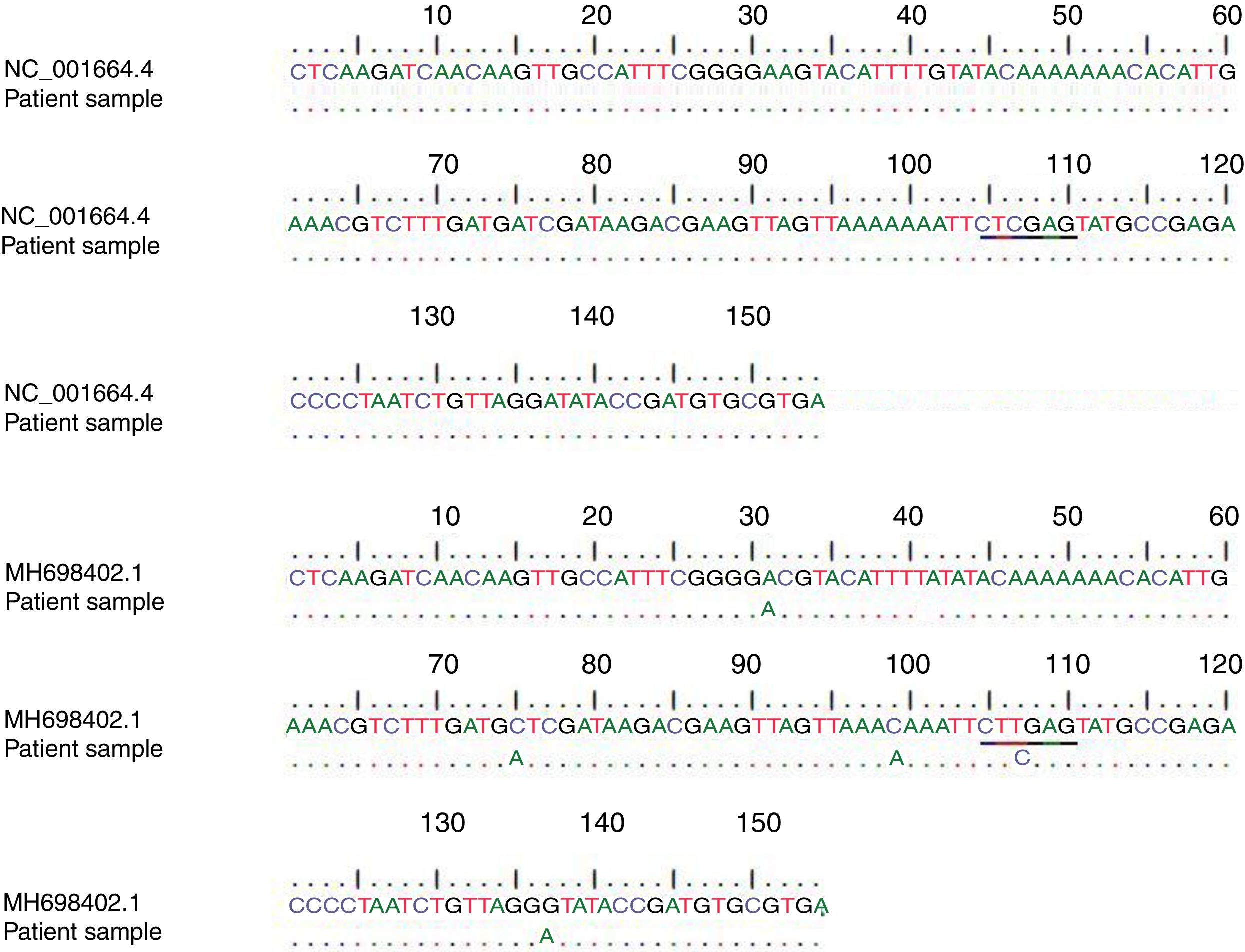
HHV-6 DNA sequencing and typingThe primers were not able to differentiate between the two types on the basis of the size of the PCR product. For discriminating between HHV-6A and HHV-6B, restriction endonuclease XhoI and nucleotide sequences were used. In all samples positive for HHV-6 DNA detected in patients either with HT or non-autoimmune thyroiditis diseases, where accurate typing result was achieved, the virus was HHV-6 type A.
In addition, the obtained sequences were subjected to a BLAST search against the GenBank database for the HHV-6A (accession no. NC_001664.4) and HHV-6B (GenBank accession no. MH698402.1) (Fig. 1). A type was assigned to each sample based on the homology of our sequence data with published sequences in GenBank as shown in Fig. 1.
Discussion and HHV-6B (MH698402.1) sequences in the GenBank database. Underlined letters indicate the position of the restriction site sequence.")
Among different environmental factors associated with autoimmune thyroiditis disease (AITD), viral infections are believed to trigger autoimmunity and may act alone or in concert with other environmental factors.21,22 Herpesviruses, in particular, have been involved in the development or exacerbation of disease.22–24 Much attention has been paid to infection with HHV-6A and AITD.
The important role of HHV-6 infection in association with autoimmune HT has generally been considered in a recent study.13 The virus was detected in 82% fine needle aspirations from HT patients. However, HHV-6 DNA was found only in 10% of the samples from patients with benign thyroid lesions. Interestingly, all samples that tested positive for HHV-6 DNA were found to be HHV-6A type, indicating that HT possibly is associated with HHV-6A infection and not with the more common HHV-6B. The evidence associated with infected thyroid cells with HHV-6 indicates that they are susceptible to natural killer cells. Therefore, the involvement of the virus in the induction of autoimmunity in HT would be expected.
In addition, detection of HHV-6 DNA, mRNA, and antigen in tissue samples of patients with autoimmune thyroiditis indicated the association of HHV-6 infection with this autoimmune disease.13,25 However, the genotype of HHV-6 had not been determined. Yet, detection of HHV-6 DNA in serum samples of patients with HT and non-autoimmune thyroiditis and in patients with euthyroid diseases had not been reported.
In our study, using a sensitive nested PCR, 38% of patients with HT were positive for HHV-6 DNA in serum samples, whereas in patients with non-autoimmune thyroid disorders only 8% was found, and in none from samples of euthyroid subjects. In terms of viral DNA present in serum samples, there was a significant difference between HT and control groups (p=0.001). HHV-6 DNA was not detected in serum samples from euthyroid patients (normal thyroid gland function with positive HHV-6 antibody). However, HHV-6 DNA was detected in serum samples from patients with autoimmune HT and in patients with non-autoimmune thyroid disorders in which anti-HHV-6 antibody was positive. Although the mean age of patients with non-autoimmune thyroiditis was higher than the other two groups, it had no influence on laboratory and HHV-6 results.
Interestingly, the variant analysis performed by XhoI restriction enzyme and DNA sequence analysis of HHV-6 DNA positive subjects indicates that all samples harbored HHV-6A, suggesting an association between thyroid diseases, in particular, HT to this virus.
As mentioned, HHV-6A/B has a tropism to thyrocytes which are permissive to virus replication.13 CD46 (also known as membrane cofactor protein or MCP) has been shown as the cellular receptor for HHV-6A entry,26 whereas CD134, a member of the TNF receptor superfamily, is the specific receptor that allows HHV-6B entry into cells.18 On the other hand, CD46 are expressed strongly on the cell surface of almost all benign and malignant thyroid follicular cells.27 Although HHV-6A/B share 90% identity in their nucleic acid sequence, they show distinct pathogenesis and cell tropism.
Some studies have reported the presence of HHV-6 DNA in PBMCs of healthy individuals.28–30 But, the detection of HHV-6 DNA by PCR in PBMCs, the site where the virus establishes latency, does not discriminate between active and latent/chronic persistent infection. However, viral load is a preferred indicator of active infection.31
Alternatively, detection of HHV-6 DNA by PCR in the serum sample is a useful indicator of active infection. However, HHV-6 DNA is not always present in plasma when the virus is active in a distant lymphoid tissue or organ.32
Even though there was no significant correlation between the presence of anti-HHV-6 antibody in serum samples among our experimental groups (p=0.3), the percentage of infected patients with HT and non-autoimmune thyroiditis diseases positive for HHV-6 antibody was higher than among normal subjects. Overall, 67% of HT patients with positive anti-HHV-6 antibody harbored HHV-6 DNA variant A in serum samples.
To clarify the association and possible role of HHV-6 as a trigger of HT, further studies are required. Several mechanisms have been suggested in which HHV-6 infections might induce an autoimmune response. However, quantification of HHV-6 viral loads in whole blood or serum samples of HT patients in association with measuring of anti-TPO antibody titer will support the role of HHV-6 active infection as a potential trigger of thyroid disease.
ConclusionHHV-6A DNA was detected in serum samples of patients with Hashimoto's thyroiditis but not in serum samples of euthyroid patients, supporting the association of HHV-6A infection and disease progression. In addition, the results of this study support other findings that HHV-6A infection might contribute to Hashimoto's thyroiditis development.
Compliance with ethical standardsEthical statementThe study was approved by the Ethics Committee of Shiraz University of Medical Sciences and has therefore been performed in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki and its later amendments.
Informed consentInformed consent was obtained from all individual participants included in the study.
Conflicts of interestThe authors declare no conflicts of interest.
Funding sourcesThis research was supported by Shiraz University of Medical Sciences grant number 1396-01-45-14603. There was no further funding from other sources to report for this submission.
The authors wish to thank Ms. Kristin Loomis, president & executive director at the HHV-6 Foundation for her invaluable assistance in editing this manuscript.
